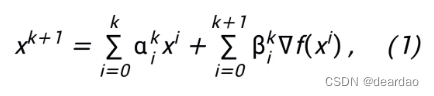文献介绍

「文献题目」 Whole-cortex in situ sequencing reveals input-dependent area identity
「研究团队」 Anthony M. Zador(美国冷泉港实验室)
「发表时间」 2024-04-24
「发表期刊」 Nature
「影响因子」 64.8
「DOI」 10.1038/s41586-024-07221-6
摘要
大脑皮层由具有不同基因表达的神经元类型组成,这些神经元被组织成专门的皮层区域。这些区域各自具有特有的细胞结构、连接性和神经元活动,并连接到模块化网络中。然而,目前尚不清楚这些空间组织是否反映在神经元转录组特征中以及这些特征在发育过程中是如何建立的。在这里,作者使用 BARseq(一种高通量原位测序技术)以细胞分辨率询问 1030 万个细胞中 104 个细胞类型标记基因的表达,其中包括 9 个小鼠前脑半球的 4,194,658 个皮层神经元。单个神经元基因表达的从头聚类揭示了与之前的单细胞 RNA 测序研究一致的转录组类型。转录组类型的组成可以高度预测皮层区域的身份。此外,具有相似转录组类型组成的区域(将其定义为皮层模块)与高度连接的区域重叠,这表明相同的模块组织反映在转录组特征和连接性中。为了探索皮层神经元的转录组学特征如何依赖于发育,作者评估了新生小鼠双目摘除后的细胞类型分布。值得注意的是,双目摘除导致视觉区域的细胞类型组成谱向同一模块内的邻近皮层区域转移,这表明外周输入使皮层模块内区域的不同转录组特性变得更加清晰。凭借 BARseq 的高通量、低成本和可重复性,作者的研究为使用大规模原位测序揭示全脑分子结构并了解其发展提供了原理证明。
研究结果
1. BARseq 绘制全脑基因表达
最近的单细胞转录组学研究使用不同的命名法来指代跨层次的细胞类型。为了避免混淆,作者首先定义了细胞类型命名法。最高层次,即 H1 型,将神经元分为兴奋性神经元、抑制性神经元和其他细胞;这个级别是许多研究中的“class”级别。在每种 H1 类型中,作者将神经元细分为 H2 类型,有时称为“subclasses”。皮层兴奋性神经元分为九种 H2 类型,大多数皮层区域共享这些类型。该分类细化了传统的基于投射的端脑内 (IT)/锥体束 (PT)/皮质丘脑 (CT) 神经元分类如下:PT 和 CT 神经元分别对应于 L5 端脑外神经元 (ET) 和 L6 CT 神经元,而 IT 神经元细分为 L2/3 IT、L4/5 IT、L5 IT、L6 IT、NP(邻近投射神经元)、Car3 和 L6b。这种划分遵循最近的单细胞 RNA 测序研究,但不同于 IT/PT/CT 神经元的经典三重分类。每个 H2 类型可以进一步分为 H3 类型(一些研究中的“cluster”或“type”级别)。先前的报告表明,H1 和 H2 类型在大多数皮层区域中基本相同,但许多基因的表达在发育过程中和成年中都局限于皮层的特定部分。H3 水平的 clusters 似乎富含来自皮质不同部分的神经元,但神经元群在皮质区域中这种更高粒度的详细分布仍不清楚。
为了评估整个皮层神经元群的分布,作者首先通过应用 BARseq 来询问覆盖一只动物整个前脑的 40 个半脑冠状切片中 104 个细胞类型标记基因(Supplementary Table 1)的表达,生成了一个试点数据集(Fig. 1a,b)。作者应用了之前用于解析运动皮层中兴奋性神经元类型的相同方法(Supplementary Note 1, Fig. 1c and Extended Data Fig. 1a–f 显示了标记基因选择和总体策略),并发现 2,167,762 个细胞跨越整个半球。在质量控制去除 reads 数不足的细胞(每个细胞 20 个 reads,每个细胞至少 5 个基因)后得到 1,259,256 个细胞(Supplementary Methods),平均每个细胞有 60 个独特 reads 和 27 个基因(Extended Data Fig. 1g,h)。在大体解剖水平上,许多基因在主要大脑结构和皮层中存在差异表达(Fig. 1a)。这些表达模式与 Allen Brain Atlas 中的原位杂交模式一致(Extended Data Fig. 1i and Supplementary Note 1)。因此,作者的试点数据集概括了已知的基因表达空间分布。
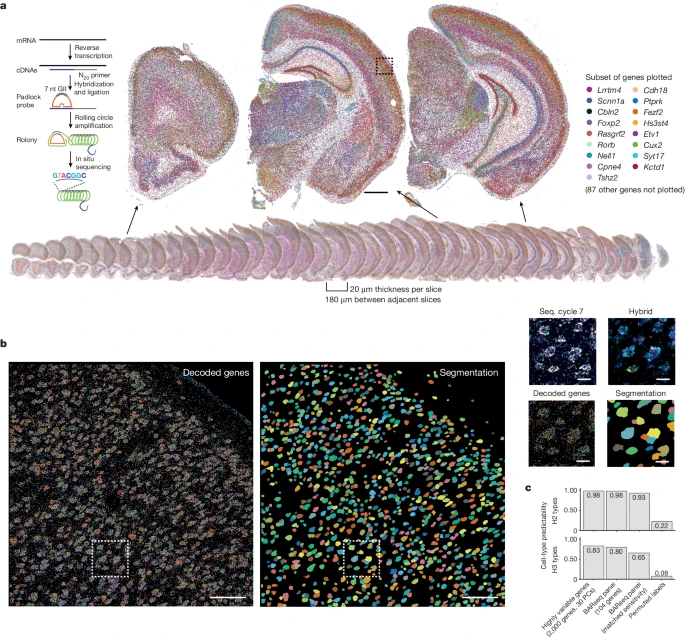
a.图像显示所有 40 个切片的 mRNA reads(底部)和三个代表性切片的特写图像(顶部)。为了清楚起见,仅绘制了 104 个基因中的 17 个(如右侧所示)。左侧插图显示了使用 BARseq 检测 mRNA 的图示。
b.从对应于 a 中的虚线框的代表性成像图块(跨 40 个切片的 4,385 个图块中)解码的基因(左)和细胞分割(中)。右侧,该区域的特写图像显示了最后一个测序周期、杂交周期、解码的基因和细胞分割。
c.使用完整转录组、top 主成分 (PCs) 和 104-gene-panel(带或不带子采样)的单细胞 cluster 分配性能,以匹配 BARseq 对 H2(顶部)和 H3(底部)clusters 的敏感性。比例尺,1mm (a),100 μm 用于全块图像 (b),10μm 用于方框区域 (b)。cDNA,互补 DNA。
2. BARseq 区分神经元类型
接下来,作者根据试点数据集中的单细胞基因表达,通过从头和层次聚类来识别神经元的转录组类型(Fig. 2a and Supplementary Methods)。将所有细胞聚类得到 24 个 clusters,然后根据 Slc17a7 和 Gad1 的表达将其组合成三种 H1 类型(642,340 个兴奋性神经元、427,939 个抑制性神经元和 188,977 个其他细胞)(Extended Data Fig. 2a and Supplementary Methods)。在这 120 万个细胞中,517,428 个位于皮层,是作者分析的重点。根据同时表达 Slc17a7 和 Gad1 的兴奋性神经元的比例,作者估计两个相邻细胞合并(即双细胞率)的分割错误概率为 5-7%(Extended Data Fig. 2b,c and Supplementary Note 2)。这 24 个 clusters 包含三种 H1 类型,很大程度上对应于大脑中的粗略解剖结构(Fig. 2b)。例如,不同的 clusters 富集在丘脑的外侧和腹侧组、层内核、上丘脑、杏仁核的内侧、基底外侧和外侧核、纹状体和苍白球(Fig. 2b)。这些结果概括了在全脑 scRNA-seq 研究中观察到的解剖学定义的大脑结构中转录组类型的明显区别。
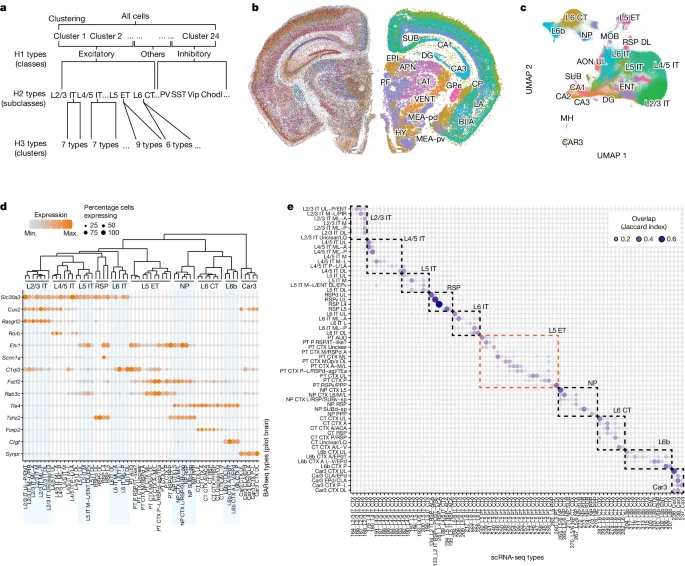
a.层次聚类的工作流程。
b.代表性切片中的基因表达(左)和 H1 clusters(右)。由 H1 clusters 区分的主要解剖分区被标记为:SUB, subiculum; DG, dentate gyrus; CP, caudate putamen; GPe, globus pallidus, external segment; LA, lateral amygdala; BLA, basolateral amygdala; MEA-pd (pv), medial amygdalar nucleus, posterodorsal (posteroventral); EPI, epithalamus; APN, anterior pretectal nucleus; LAT, lateral group of the dorsal thalamus; VENT, ventral group of the dorsal thalamus; PF, parafascicular nucleus; HY, hypothalamus。
c.兴奋性神经元基因表达的 UMAP 图,按 H2 类型着色。CA, cornu Ammonis; MOB, main olfactory bulb; MH, medial habenula; AON, anterior olfactory nucleus; RSP, retrosplenial area。
d.皮层兴奋性 H3 类型中的标记基因表达。颜色表示平均表达水平,点大小表示表达该基因的细胞分数。树状图(顶部)显示了每种 H3 类型内汇集基因表达的层次聚类。
e.BARseq H3 类型和 scRNA 细胞类型之间的重叠(Jaccard 指数)。虚线框表示父代 H2 类型。Min.,最小;max.,最大。
然后,作者将兴奋性和抑制性神经元分别重新聚类为 H2 类型(Fig. 2a,c and Extended Data Fig. 2d)以提高聚类的分辨率。在这个水平上,作者恢复了主要的抑制性神经元亚类(Pvalb、Sst、Vip/Sncg、Meis2-like 和 Lamp5),以及整个皮层共享的所有兴奋性亚类(L2/3 IT、L4/5 IT、L5 IT、L6 IT 、L5 ET、L6 CT、NP、Car3 和 L6b)以及先前皮层 scRNA-seq 数据集中观察到的针对内侧皮质 (RSP) 的兴奋性子类。 H2 类型表达已知的细胞类型标记和其他高度差异表达的基因(Fig. 2d)。例如,Cux2 主要在浅层 IT 和 Car3 神经元中表达,Fezf2 在 NP 和 L5 ET 神经元中表达,Foxp2 特别在 L6 CT 神经元中表达(Supplementary Note 3 提供了详细描述)。尽管作者分两批生成了完整的 40-切片 数据集(Supplementary Methods),但没有观察到强烈的批次效应,正如 UMAP 图中来自两批不同切片的兴奋性神经元的混合所证明的那样(Extended Data Fig. 2e)。因此,H2 类型以中等粒度概括了先前 scRNA-seq 数据集中识别的已知神经元类型。
然后,作者将每种兴奋性 H2 类型重新聚类为 H3 类型(Fig. 2a)。为了量化 H3 类型与之前 scRNA-seq 研究中确定的参考转录组类型的对应程度,作者使用基于 k 最近邻的方法将每个 H3 类型与参考文献中记录的叶子级 clusters 进行匹配 (Supplementary Methods)。作者发现皮层 H2 类型与 scRNA-seq 数据中的亚类水平细胞类型具有一一对应的关系(Fig. 2e)。在每个 H2 类型中,H3 类型差异地映射到 scRNA-seq 数据中叶子级 cluster 的单个或小子集(Fig. 2e; Extended Data Fig. 2f 显示了皮层外部 clusters 的匹配)。H2 和 H3 类型都沿着皮层深度以有序的方式组织,概括了皮层兴奋性神经元的层状组织(Extended Data Fig. 2g,h and Supplementary Note 3)。在粗空间分辨率下,H3 类型也在皮层区域中被发现,类似于之前的 scRNA-seq 数据集中的匹配 clusters(Extended Data Fig. 2i–k and Extended Data Fig. 3)。例如,H3 型 PT AUD 及其相应的 scRNA-seq cluster(242_L5_PT CTX)均富集于外侧皮质区(TEa-PERI-ECT)和听觉皮层(AUD),而 H3 型 PT CTX P 及其相应的 scRNA-seq clusters(245_L5_PT CTX 和 259_L5_PT CTX)在视觉皮层中富集。因此,这些结果表明,作者的试点数据集解析了皮质兴奋性神经元的细粒度转录组类型,这与之前的 scRNA-seq 数据集一致,并概括了它们的区域和层状分布。作者的数据集的高分辨率和皮层范围现在使我们能够以微米级分辨率解析基因表达的空间富集和神经元亚群在整个皮层的分布。
3. 整个皮层的基因表达模式
整个皮层的基因表达差异很大,但大多数皮层区域在很大程度上共享相同的兴奋性神经元 H2 类型或亚类。因此,尚不清楚神经元亚群组织的差异如何导致区域特异性基因表达。三个变异来源可能导致不同区域的基因表达差异(Fig. 3a)。首先,H2 类型的组成可能会导致整个皮层基因表达的差异(Fig. 3a(左),细胞类型组成模型)。例如,H2-type X 与 H2-type Y 的比率可能在视觉皮层中较高,但在运动皮层中较低,因此在 X 中表达高于 Y 的基因在视觉皮层中表达较高。其次,无论 H2 类型如何,某些基因的表达可能会在空间上发生变化,也就是说,它们在多种 H2 类型中跨空间一致变化(Fig. 3a(中),空间梯度模型)。在该模型中,基因 A 在 X 型和 Y 型中在视觉中的表达可能比在运动皮层中的表达更高。最后,一些基因的表达可能以 H2 特异性方式跨空间变化(Fig. 3a(右),区域专用细胞类型模型)。例如,在 H2 X 型中,基因 A 在视觉皮层中的表达可能比在运动皮层中更高,但在 H2 Y 型中则不然。
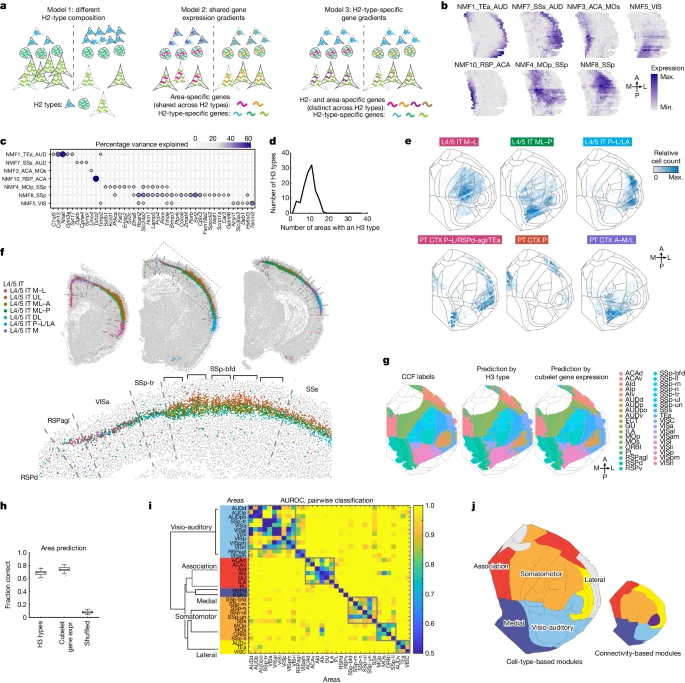
a.皮层区域差异基因表达的三种模型。H2 型由细胞形状表示;冷色调形状表示 H2 型特异性基因;暖色调形状表示模型 2 中的区域特异性基因和模型 3 中的 H3 型特异性基因。
b.绘制在皮质平面图上的选定空间变异 NMF 模块的表达。
c.每个 NMF 模块中所选标记基因的表达。
d.直方图显示检测到 H3 类型的区域数量(共 37 个):如果某个区域包含至少 3% 的 H3 类型,则认为该区域中存在 H3 类型。
e.在皮质平面图上绘制的示例 L4/5 IT 和 L5 ET H3 类型在皮质上的空间分布。颜色表示每个立方体中的相对细胞计数;灰线描绘了皮质区域。
f.示例冠状切片中 L4/5 IT H3 类型的分布。虚线表示 CCF 中的区域边界。虚线框的放大视图显示在右侧。括号表示桶状皮层中的桶。
g.CCF 中定义的皮质区域(左)以及通过 H3 类型(中)和立方体基因表达(右)预测的皮质区域。 h. 使用 H3 型组成、立方体基因表达和混洗对照正确预测立方体的分数。每个框显示 n = 100 次重采样试验的性能。方框显示中位数,四分位数和须线表示排除异常值后的范围。点表示异常值。
i.矩阵显示皮质区域组合之间成对分类的 AUROC。区域按模块排序,模块在左侧用颜色编码。利用H3 型组成的相似性计算树状图;聚类是根据矩阵获得的,并显示在灰色边框框中。
j.由基于细胞类型的模块(左)和 Harris et al. 识别的基于连接的模块(右)着色的皮质平面图。
为了确定每个来源对跨区域基因表达变化的贡献,作者将每个冠状切片上的皮质离散成 20 个空间 bins(Supplementary Methods and Extended Data Fig. 4a)。然后,作者使用单向方差分析(Extended Data Fig. 4b,c and Supplementary Methods)评估了跨 bins 的大量基因表达的变异有多少可以通过 H2 或 H3 类型的空间或组成来解释。作者发现所有三个模型都会导致基因表达的空间变异,并且对变异贡献最大的模型因基因而异(Extended Data Fig. 4b–d and Supplementary Note 4)。由于许多基因的空间模式相似,作者试图使用非负矩阵分解 (NMF) 提取基因和 H2 类型之间共享的基本空间成分(Supplementary Note 4 and Extended Data Fig. 4e,f)。作者发现,大多数 NMF 成分并不是沿主要空间轴呈宽梯度分布,而是集中在功能相关且高度互连的区域(Fig. 3b and Extended Data Fig. 4g)。例如,NMF5 主要存在于视觉区域,而 NMF8 主要存在于体感区域。其他 NMF 模块,包括 NMF1(内侧区域)和 NMF10(外侧区域),存在于功能不同但高度互连的区域组合中。空间变异基因通常仅与一种或两种成分密切相关(Fig. 3c and Extended Data Figs. 4h and 5),并且这种关联概括了这些基因的已知空间模式。例如,Tenm3 主要在后感觉区域表达,包括视觉皮层、听觉皮层和部分体感皮层(Extended Data Fig. 4d,底部);Tenm3 与 NMF5 密切相关(Fig. 3c),NMF5 也在相同的区域组中表达(Fig. 3b)。因此,基因表达沿着相互关联的区域组变化,这表明基因表达和跨区域的皮质内连接之间存在有趣的联系。
4. 细胞类型定义的皮层模块
在控制了 H2 类型(而非 H3 类型)组成的变异性后,获得了空间变化的 NMF 模块。因此,作者假设这些模块反映了皮层区域 H3 类型组成的差异。与这一假设一致,每种 H3 类型都富含 NMF 模块的一小部分,并且 H3 类型也与其相应的 NMF 模块在空间上重叠(Extended Data Figs. 6 and 7a,b, Supplementary Methods and Supplementary Note 5)。为了进一步评估 H3 类型的区域分布,作者将每个冠状切片上的皮层重新离散成沿着所有切片的中外侧轴宽度相似的“cubelets”(Supplementary Methods and Extended Data Fig. 4a)。这些 cubelets 具有相似的物理尺寸,并且在中外侧轴上比先前分析中使用的空间 bins 更窄;这种更高的横向分辨率使得更容易将 cubelets 分配到各个皮层区域。作者发现 H3 类型由多个皮层区域共享,并且不特定于任何单个区域(每种 H3 类型发现于 6 到 12 个区域之间,median ± 1 s.d.,Fig. 3d; Fig. 3e 显示了示例 H3 类型的分布)。因此,相邻皮层区域的独特性不能简单地通过区域特异性 H3 类型的存在或不存在来解释。然而,作者注意到,H3 类型的成分分布经常在 Allen 通用坐标框架 v.3 (CCF) 中定义的区域边界附近突然发生变化(Fig. 3f and Extended Data Fig. 7c,d)。最显着的变化发生在外侧和内侧区域,这与 scRNA-seq 数据一致。在背外侧皮层内,虽然邻近的皮层区域有时共享 H3 类型组,但它们的比例通常在区域边界处或附近发生变化。使用每个 cubelets 中的基因表达或 H3 类型的组成,我们可以准确预测 cubelets 的位置和皮层区域标签(使用基因表达正确率为 75%,使用 H3 类型组成正确率为 69%,而 shuffled 对照中为 8%;Supplementary Note 5, Fig. 3g,h and Extended Data Fig. 7e–i)。因此,cubelets 基因表达和 H3 型组成都可以高度预测沿皮质切面的位置和皮质区域的身份。
接下来,作者根据 H3 型组成的区分程度来评估皮层区域的相似性和模块化(Fig. 3i and Supplementary Methods)。简而言之,作者基于使用 H3 类型组合对皮层区域进行成对区分的程度构建了皮层区域之间的距离矩阵,然后对距离矩阵进行了 Louvain 聚类。作者确定了六个 clusters,每个 cluster 都包含多个区域(Fig. 3i,灰色方框);其中包括对应于视觉听觉区域的两个 clusters,以及对应于关联区域、体感皮层、运动皮层和侧向区域的各一个 cluster。这种模块化组织对于 CCF 注册中的小错误具有鲁棒性(Extended Data Fig. 7j and Supplementary Methods)。基于 H3 型组成的相似性,作者进一步将这些 clusters 与不与任何其他区域聚类的单个区域(PL、RSPd 和 RSPv)组合成皮层模块。这些模块主要分别包括视觉听觉、躯体运动、关联、内侧和外侧区域(Fig. 3i)。值得注意的是,这些基于细胞类型的模块与高度连接的皮层模块(基于连接的模块)基本一致(Fig. 3j)。因此,高度互连的皮层区域共享 H3 类型组,因此具有特征转录组显著性。
5. 细胞类型对双目摘除具有鲁棒性
转录组类型、区域和模块反映了不同尺度的皮层组织,表明它们可能是通过不同的发育机制产生的。作为了解不同尺度的皮层组织发育过程的第一步,作者应用 BARseq 来研究出生后外周感觉输入的去除如何改变皮层转录组类型的组织。丘脑皮层投射在塑造皮层区域的身份和边界方面发挥着核心作用,出生后视觉输入的丧失会影响 VISp 和其他区域的基因表达。然而,外周输入如何塑造皮层神经元类型和皮层区域的特征性细胞类型组成特征尚不清楚。例如,基因表达的改变可能会产生正常大脑中看不到的新细胞类型;或者,它可以富集或丢失现有的细胞类型(Fig. 4a)。由于 BARseq 具有成本效益且具有高通量,因此它特别适合在许多动物的大脑范围内询问神经元基因表达和细胞类型组成谱的变化,无论是否存在发育扰动。
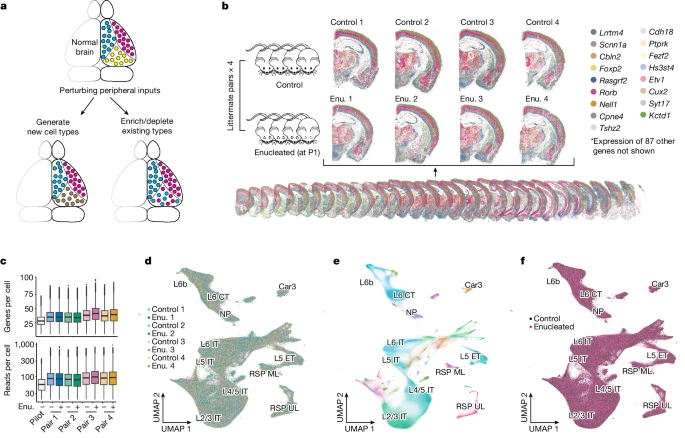
a.出生后去除外周感觉输入可能产生的影响模型,包括新细胞类型的产生(左)和/或现有细胞类型的富集和丢失(右)。
b.从四对同窝小鼠中收集了全脑转录组数据:每对中,一只小鼠在 P1 时被摘除双目(Enu.),另一只是假对照(左,n = 8 只动物)。显示了来自一个大脑的 32 个切片的代表性堆栈(底部)和来自所有八个大脑的匹配冠状切片的特写图像(顶部)。为了清楚起见,仅绘制了 104 个基因中的 17 个(如右侧所示)。
c.试验数据集以及去核和对照同窝小鼠的每个细胞的基因和 read 计数(按顺序绘制,n=0.6、1.0、1.0、1.1、1.3、1.2、1.1、1.2 和 1.1 百万个生物独立细胞)。方框表示中位数以及第一和第三四分位数;晶须从每个边界延伸至最极端值,最多可达四分位数间距的 1.5 倍;剩余的点单独绘制。
d-f.所有八只动物兴奋性神经元基因表达的 UMAP 图。神经元按动物 (d)、H2 类型 (e) 和条件(双目摘除或对照,f)进行颜色编码。标签仅显示皮层中的 H2 类型。
在出生后第 1 天对四只小鼠进行了双目摘除术,并在出生后第 28 天收集了它们的大脑,以及四只匹配的同窝对照动物(n = 8 只动物)的大脑(Fig. 4b)。作者使用改进的显微镜进行 BARseq,与试点数据集相比,该显微镜实现了更好的数据质量和更快的数据采集(每个大脑 2.3 天;Supplementary Methods and Supplementary Note 6)。整个数据集总共包含 9.1 百万个质量控制细胞,覆盖所有八只动物的大部分前脑(Fig. 4b),每个细胞的中位数为 87 个 reads 和 37 个基因(Fig. 4c)。来自单个大脑的细胞与 UMAP 空间中其他大脑的细胞相互交错,表明批次效应最小(Fig. 4d and Extended Data Fig. 8a)。因此,作者对 H1 级别的 3,957,252 个兴奋性神经元、1,526,182 个抑制性神经元和 3,635,402 个其他细胞的串联数据进行了从头聚类。其他细胞的比例明显高于试点数据集中的比例,可能是因为数据质量的提高允许更多 read 计数较低的细胞通过质量控制并被纳入。然后,作者将兴奋性神经元重新聚类为 35 个 H2 类型(Fig. 4e)和 154 个 H3 类型,包括主要在皮层中发现的 12 个 H2 类型和 70 个 H3 类型。新数据集中的这些 H3 类型与试点数据集中的 H3 类型密切匹配(Extended Data Fig. 8b,c; Supplementary Note 6 显示了到试点数据集的映射)。值得注意的是,与对照相比,双目摘除大脑中没有 H3 类型强烈富集或丢失(Extended Data Fig. 8f; Supplementary Note 7 and Extended Data Fig. 8d–i 提供了详细分析)。尽管不能完全排除在作者的转录组分辨率下遗漏基因表达微小变化的可能性,但这些结果表明双目摘除并没有导致 H3 水平上新细胞类型的产生;相反,双目摘除的主要影响可能反映在 H3 类型成分谱的变化上。
6. 双目摘除改变细胞类型组成
确定双目摘除不会产生新的 H3 类型后,作者试图表征双目摘除引起的区域特异性 H3 类型组成的变化。作者使用类似于试验数据的方法(Supplementary Methods)将皮层划分为 cubelets。这种离散化导致每个 cubelets 大约有 270 个神经元,切片中相邻 cubelets 之间的平均距离为 181 µm。为了可视化 H3 类型组成,作者根据每个 cubelets 中 H3 类型的比例绘制了 UMAP 分析图(Fig. 5a-c)。与单神经元基因表达中不存在批次效应一致(Fig. 4d),来自所有八只动物 cubelets 在大部分区域顺利混合(Fig. 5a)。然而,按条件对 cubelets 进行颜色编码(Fig. 5b),显示出一个“island”(左),其中来自两个群体(双目摘除与对照)的 cubelets 在很大程度上被隔离。该 island 主要包含来自 VISp 和其他视觉区域的 cubelets(Fig. 5b、c,插图)。为了量化对照大脑和双目摘除大脑之间 H3 类型的组成特征的差异,作者训练了一个分类器来评估两种条件下每个皮层区域的 cubelets 的差异(Supplementary Methods)。如果双目摘除持续改变了皮层区域中 H3 类型的组成特征,那么我们期望分类器能够根据高于机会水平的 H3 类型组成来预测 cubelets 是来自对照动物还是双目摘除动物。在大多数皮层区域,分类器在机会水平上执行,但 VISp cubelets 对条件具有高度预测性(Fig. 5d)。两个较高的视觉区域(VISpm 和 VISl)和一个非视觉区域(SSp-ll)的预测也高于机会水平,尽管预测能力要低得多。因此,双目摘除很大程度上影响了视觉区域内 H3 类型的相对组成。
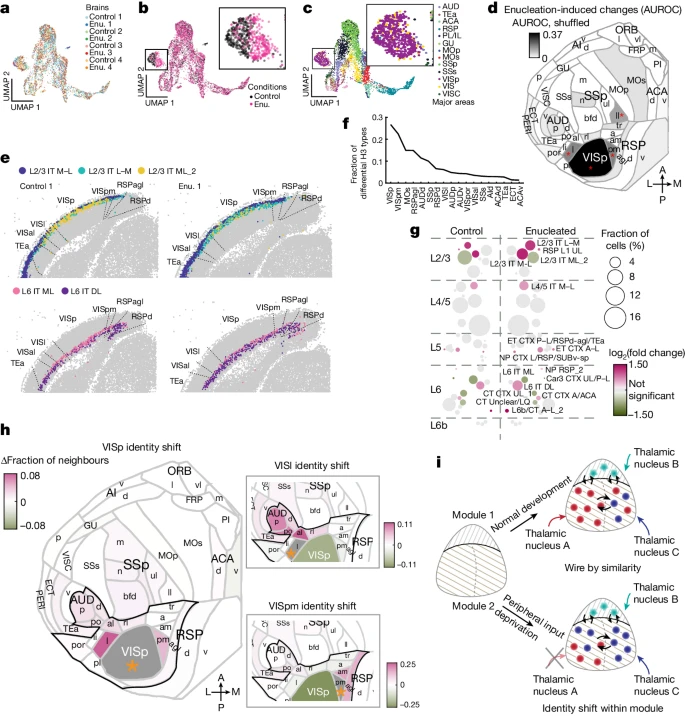
a-c.来自所有八个大脑的 cubelets 的 H3 类型组成的 UMAP 图,按动物 (a)、实验条件 (b) 和皮层区域 (c) 进行颜色编码。插图显示了方框区域的放大视图。
d.平面图显示对照和双目摘除大脑(AUROC)之间的区域有多么不同。颜色表示有或没有混洗大脑条件的分数之间的 ΔAUROC。红色星号表示错误发现率低于 0.05。
e.来自一对同窝小鼠的代表性切片,显示双目摘除后选定的 L2/3 IT 和 L6 IT H3 类型在 VISp 中富集或丢失。
f.每个区域中 H3 类型富集或丢失的分数。
g.VISp 中 H3 类型的分数及其双目摘除后的倍数变化。颜色表示 log(fold change),圆圈大小表示属于每种 H3 类型的 VISp 神经元的分数。
h.VISp(左)、VIS1(右上)和 VISpm(右下)的面域特性变化。颜色表示与对照相比,双目摘除大脑中富集或丢失的邻居的比例。每个图中感兴趣的区域都以灰色阴影显示,并用橙色星号表示。黑色轮廓的区域表示视觉听觉模块。
i.皮层细胞类型组织模型。皮层模块的建立独立于外周输入(左)。在正常情况下,皮层区域遵循逐条相似性(上)。模块内的区域具有相似的细胞类型并且更加互连。当外周输入被移除时(底部),受影响区域的细胞类型组成轮廓会转向同一模块内的其他区域。模块由带图案的背景表示。彩色点和星号分别表示两个模块中的细胞类型;彩色箭头表示丘脑输入;皮层内的黑色箭头表示连接性。
双目摘除的效果可以直接在初级视觉区域的 H3 类型分布中观察到(Fig. 5e and Extended Data Fig. 9a)。例如,在对照动物的 VISp 中发现了许多 L2/3 IT M-L_2 神经元(Fig. 5e,黄点),但在双目摘除动物的 VISp 中 L2/3 IT L–M 神经元(Fig. 5e,绿点)富集。同样,与对照动物相比,在双目摘除动物的 VISp 中发现 L6 IT DL 神经元(Fig. 5e,紫色点)的数量更高。为了系统地研究双目摘除如何影响每个区域的皮层兴奋性细胞类型的组成特征,作者使用方差分析模型寻找在双目摘除大脑中富集或丢失的 H3 类型,调整垃圾和区域效应(Supplementary Methods)。作者发现,与对照组相比,双目摘除动物整个皮层 18 个区域的 46 个 H3 类型要么过多,要么过少。VISp 具有最多的 H3 类型 (16),其组成因双目摘除而改变(Fig. 5f)。在大多数 H2 类型中都发现了受影响的 H3 类型,其中 L2/3 IT、L4/5 IT 和 L6 IT 的 H3 类型的富集或丢失最强(Fig. 5g)。有趣的是,L6b/CT A–L_2(L6 CT 和 L6b H2 类型之间的过渡类型,通常仅在侧面区域发现)在摘除后在 VISp 中也高度富集。受影响的 H3 类型保持在其特征性的层下位置(Extended Data Fig. 9b),总体变化与关键期黑暗饲养期间观察到的变化一致,但范围更广(Extended Data Fig. 9c and Supplementary Note 8)。最富集的 H3 类型,包括 L2/3 IT M–L、L2/3 IT L–M、L4/5 IT M–L、L6 IT DL 和 L6b/CT A–L_2,均在内侧和外侧区域富集在对照大脑中,包括紧邻视觉区域内侧和外侧的区域(Extended Data Fig. 9d)。因此,摘除后,VISp 中的神经元广泛转变为 H3 类型,这些类型通常富含于对照大脑的内侧和外侧区域。
7. 外周输入塑造区域身份
由于在对照动物的内侧和/或外侧区域始终发现富集的 H3 类型,作者想知道双目摘除是否也将视觉皮层的整体区域特征(由 H3 类型组成特征定义)转移到其他区域。为了检查双目摘除后区域身份如何变化,作者使用受 MetaNeighbor 启发的基于最近邻的方法来评估对照大脑和双目摘除大脑中的 cubelets 与对照大脑中其他 cubelets 的相似性(Supplementary Methods)。如果双目摘除使一个区域的成分分布向目标区域转移,那么来自被双目摘除大脑中受影响区域的 cubelets 在目标区域中将比来自对照大脑中相同区域的 cubelets 有更多的邻居。对于同窝小鼠的每对 cubelets,作者在其他三对同窝小鼠的对照大脑中找到了 20 个与 H3 类型组成最匹配的 cubelets。然后,作者计算了相似性,通过 AUROC 进行量化,根据最近邻将每个区域的 cubelets 分配给控制大脑中的区域(Extended Data Fig. 9e)。所有三个视觉区域(VISp、VISl 和 VISpm,在 Extended Data Fig. 9e 中圈出)与对照大脑中的相同区域保持高度相似(对照和双目摘除 VISp 的 AUROC 为 0.97 和 0.98,对照和双目摘除 VISl 的 AUROC 为 0.90 和 0.92,对照和双目摘除 VISpm 的 AUROC 为 0.88 和 0.94),表明尽管双目摘除引起了变化,但它们的 H3 类型组成仍然与其他区域高度不同。然而,根据一个区域的邻居比例来判断,所有三个视觉区域也转向邻近区域的身份(Fig. 5h)。例如,与来自对照大脑的那些相比,来自双目摘除大脑的 VISp cubelets 具有更高的 VIS1 和 VISpm AUROC 得分(双目摘除 cubelets 为 0.85 和 0.89,对照 cubelets 为 0.76 和 0.83;Extended Data Fig. 9e)。与观察到的高 AUROC 分数一致,来自双目摘除大脑的 VISp cubelets 在 VIS1 和 VISpm 中也有更多的邻居(Fig. 5h)。类似地,来自双目摘除大脑的 VIS1 cubelets 在听觉区域有更多邻居,来自双目摘除大脑的 VISpm cubelets 在 VISam 和 RSPagl 中有更多邻居(Fig. 5h,插图)。值得注意的是,所有三个区域都向物理上远离 VISp 且位于视觉听觉模块内的邻近区域转移(Fig. 5h 中的黑色轮廓)。为了检查这些变化是否反映了区域边界的变化或整个区域成分的变化,作者绘制了来自双目摘除大脑的每个 cubelets,并根据 VIS1 中相邻 cubelets 数量的差异对它们进行着色(Extended Data Fig. 9f,顶部)和 VISpm(Extended Data Fig. 9f,底部)。在 VISp 中,在整个区域的 cubelets 中发现了 VIS1 和 VISpm 中邻居的富集。特别是,双目摘除后在 VISpm 中有更多邻居的 cubelets(Extended Data Fig. 9f 中 VISp 中的红点,底部)似乎集中在 VISp 的中心而不是边界,表明这些区域之间的相似性变化反映了细胞类型组成的整体变化而不是区域边界的变化。因此,双目摘除将视觉区域的 H3 型成分定义的区域身份转移到视觉听觉模块内的邻近区域。
讨论
使用 BARseq,作者在九只动物中以高转录组和空间分辨率生成了兴奋性神经元转录组类型的皮层范围图。这些图不仅详细阐述了先前通过单细胞研究揭示的皮质兴奋性神经元类型的分布,而且还提供了将其他神经元特性和活动与神经元类型相关联的“锚点”。因此,作者的空间细胞类型图为理解皮层区域的结构和功能特异性提供了基础资源。作者专注于皮层,但同样的方法可以应用于任何其他具有充分设计的基因组的大脑区域。当检查大量基因时,通过重叠信号的计算分离和/或使用更新的方法优化细胞分割来克服光学拥挤可能会进一步提高准确解析单细胞基因表达的能力。
作者的结果表明,皮层区域的细胞类型组成概况反映了其在连接性研究中看到的模块化组织:高度互连的皮层区域也具有相似的 H3 类型(Fig. 5i,顶部)。这种“wire-by-similarity”关系并不是在整个皮层范围内观察到的细胞类型特异性连接的微不足道的结果,因为相同类型的皮层神经元不一定是高度连接的(例如,Sst 神经元)。因此,wire-by-similarity 并不描述单个神经元类型的连接性,而是反映了大的大脑区域(即皮层内的区域)内的分裂在细胞类型和连接性方面如何相互关联。未来的研究使用 BARseq 从多个皮层区域和多个发育时间点以细胞分辨率绘制神经元类型的投射,可以帮助解决 wire-by-similarity 组织的单细胞基础问题。
单细胞分辨率、高转录组分辨率和跨多个皮层区域的广泛询问相结合,使作者能够详细描述在去除外周感觉输入后基因表达和细胞类型组成谱如何变化。总体而言,双目摘除的影响表明外周活动改善了皮层区域的细胞类型组成特征。双目摘除影响所有层的 IT 神经元以及 L6b/CT 神经元,其群体比受黑暗饲养影响的 L2/3 IT 神经元更广泛(Fig. 5g)。然而,正如 Chou et al. 在丘脑皮质轴突基因消融后观察到的那样,双目摘除并没有完全消除初级和次级视觉区域之间的区别(Supplementary Note)。因此,与之前的研究一起,作者的结果提出了一个一致的模型:丘脑皮层轴突建立的物理连接是定义初级视觉皮层所必需的,而外围活动则使初级视觉皮层和皮层模块内邻近高级视觉区域的细胞类型组成变得更加清晰(Fig. 5i,底部)。
BARseq 以其高通量(每个大脑在一台显微镜上大约 2.3 天)、低成本(每个大脑大约 2,000 美元)和高重复性而在空间转录组学方法中脱颖而出。这些特征使得比较许多动物的全脑空间基因表达成为可能,从而提供了一条超越单一参考大脑图谱,迈向捕捉种群多样性的“泛转录组”图谱的途径。此外,将多个个体的询问与扰动结合起来可以发现因果关系。尽管作者研究了发育扰动的影响,但同样的方法也可用于神经精神疾病模型、衰老研究、跨物种比较和其他实验扰动。作者基于 BARseq 的方法可广泛应用于将全脑网络级动态与单个神经元基因表达的详细变化联系起来,并建立发育过程和全脑细胞类型组织之间的因果关系。
注:本文为个人学习笔记,仅供大家参考学习,不得用于任何商业目的。如有侵权,请联系作者删除。

本文由 mdnice 多平台发布